|
 Life
Cycle of Listeria monocytogenes The
life cycle of the bacterial pathogen, Listeria
monocytogenes is summarized
in the image at the right (from
Tilney & Portnoy 1989, J
Cell Biol 109:1597-1608). As
part of normal defenses against
infection, white blood cells
(macrophages) "eat" Listeria (phagocytosis),
intending to destroy the bacterium. However, Listeria makes
specific enzymes that help it
escape the cell's "stomach" (lysosome)
into the cytoplasm. Somewhat "hidden" by
the "cloak" of the
host cell's outer membrane, Listeria evades
the immune system by growing within the
cytoplasm of host cells. Life
Cycle of Listeria monocytogenes The
life cycle of the bacterial pathogen, Listeria
monocytogenes is summarized
in the image at the right (from
Tilney & Portnoy 1989, J
Cell Biol 109:1597-1608). As
part of normal defenses against
infection, white blood cells
(macrophages) "eat" Listeria (phagocytosis),
intending to destroy the bacterium. However, Listeria makes
specific enzymes that help it
escape the cell's "stomach" (lysosome)
into the cytoplasm. Somewhat "hidden" by
the "cloak" of the
host cell's outer membrane, Listeria evades
the immune system by growing within the
cytoplasm of host cells.
Once
within the cytoplasm of the host
cell, Listeria quickly
assembles proteins from the host
cell to form rocket-like tails
containing F-actin. These
F-actin tails propel bacteria
through the cytoplasm (see QuickTime
movie (3.8Mb) of F-actin tail
propelling bacterium inside a
host cell). When bacteria
encounter the cell's outer membrane,
they deform the membrane and
attempt to infect neighboring
cells. During the protrusion
of Listeria, bacteria
maintain the protection of original
host cell's outer membrane. Long
before the original host cell
fills with bacteria and bursts,
adjacent cells have already been
infected.
The generation of a
rocket-like F-actin tail and
subsequent propulsion of bacteria
is essential to Listeria's
pathogenicity. Only one
bacterial protein (ActA) is required
to generate rocket tails. Genetic
deletion of this protein nearly
blocks the virulence of this
bacterium.
Relevance
to Other Pathogens and Cell Motility Other
pathogens also generate propulsive
F-actin tails. These include Shigella (bacterial
dysentery), Rickettsia (Rocky
Mountain spotted fevers), cowpox
(vaccinia) virus,and, presumably,
smallpox virus. By analogy
to Listeria, all these
pathogens probably use their
F-actin rocket-like tails for
motility and to infect neighboring
cells. Hence, the rocket-tail
propulsion would be critical
for the virulence of these pathogens.
Not
only an interesting bacterial
pathogen, Listeria is
a model system for understanding
cell motility. The same
host proteins recruited by Listeria are
also used by the host cell for
it's own motile and protrusive
processes. When cells crawl
(e.g. activated white blood cells),
they must protrude their leading
edge prior to gripping and pulling. Similarly,
when cells "eat" (phagocytose),
cells must locally protrude to
engulf particles (see image above;
see QuickTime
movie of white blood cell "hunting
down" a bacterium, 9Mb). Finally,
when neuronal cells are extending
axons, the leading edge of the
axon protrudes finger-like filopodia
to "taste" for guidance
cues that direct neurite extension
(see QuickTime
movie of neuronal filopodia,
2Mb). Activated by
the similar molecular processes,
all of these processes require
actin polymerization by the host
cell to protrude. Because Listeria generates "stripped-down",
unregulated motility, it helps
focus investigators on the key
host proteins required for all
these motilities. So far,
only proteins required to regulate
actin polymerization into F-actin
are believed necessary for Listeria motility
(Loisel et al. 1999, Nature 401:613-615).
 How
can mere polymerization push? Despite
its widespread acceptance, it
seems paradoxical that actin
polymerization alone can generate
forces. To illustrate this
paradox, consider the cartoon
at the right and its companion
animation (either Flash,
5Kb or QuickTime,
7Kb). Think of actin
monomers as books being stacked
onto a bookshelf. When
reaching the bookend, there's
not enough room for the next
book. If we were to manually
slide the bookend to make room
for the next book, our action
of moving the bookend generated
the force; stacking books only
stabilized the new position of
the bookend. From this
macroscopic analogy, polymerization
alone could never generate forces. How
can mere polymerization push? Despite
its widespread acceptance, it
seems paradoxical that actin
polymerization alone can generate
forces. To illustrate this
paradox, consider the cartoon
at the right and its companion
animation (either Flash,
5Kb or QuickTime,
7Kb). Think of actin
monomers as books being stacked
onto a bookshelf. When
reaching the bookend, there's
not enough room for the next
book. If we were to manually
slide the bookend to make room
for the next book, our action
of moving the bookend generated
the force; stacking books only
stabilized the new position of
the bookend. From this
macroscopic analogy, polymerization
alone could never generate forces.
 This
apparent paradox can be resolved
because nothing is truly stationary
at the microscopic size scale. As
shown in the cartoon at right
with its companion animation
(either Flash,
8Kb or QuickTime,
9Kb), heat (even at room
temperature) causes all objects
to "wiggle" with Brownian
motion. The magnitude of
Brownian motions are generally
inversely related to size; hence,
microscopic objects can "wiggle" a
lot. In the macroscopic
analogy, the bookend intrinsically
wiggles more than the size of
each book, and when it wiggles
far enough, polymerization would "jam" the
next book into the gap on the
bookshelf. Each inserted
book prevents any backsliding
and "ratchets" the
bookend forward. Called
the "Brownian ratchet" by
George Oster and colleagues,
this type of model is very elegant
and amenable to analytical exploration
with a minimum of parameters
(Peskin, Odell & Oster 1993, Biophys
J 65:316-324). To
distinguish it from later Brownian
ratchet models, it is often called
the "classic Brownian
ratchet" model. This
apparent paradox can be resolved
because nothing is truly stationary
at the microscopic size scale. As
shown in the cartoon at right
with its companion animation
(either Flash,
8Kb or QuickTime,
9Kb), heat (even at room
temperature) causes all objects
to "wiggle" with Brownian
motion. The magnitude of
Brownian motions are generally
inversely related to size; hence,
microscopic objects can "wiggle" a
lot. In the macroscopic
analogy, the bookend intrinsically
wiggles more than the size of
each book, and when it wiggles
far enough, polymerization would "jam" the
next book into the gap on the
bookshelf. Each inserted
book prevents any backsliding
and "ratchets" the
bookend forward. Called
the "Brownian ratchet" by
George Oster and colleagues,
this type of model is very elegant
and amenable to analytical exploration
with a minimum of parameters
(Peskin, Odell & Oster 1993, Biophys
J 65:316-324). To
distinguish it from later Brownian
ratchet models, it is often called
the "classic Brownian
ratchet" model.
 To
give the classic
Brownian ratchet some
molecular details, consider the
cartoon at right and its companion
animation (either Flash,
7Kb or QuickTime,
9Kb). Actin monomers
are 5.4 nm in diameter and F-actin
filaments are composed of two
intertwined protofilaments that
are staggered by half a monomer
(2.7 nm). As suggested
by the cartoon and its animation, Listeria must "wiggle" at
least 2.7 nm to allow intercalation
of actin monomers. The
magnitude of the "wiggles" are
inversely proportional
to the size of the bacterium
and to the viscosity of its environment. Because
diffusion is time-dependent,
the longer you wait, the larger
the magnitude of Brownian motions
and intercalation eventually
occurs. However, the high
speed of Listeria motility
implies that bacteria diffuse
very readily. Rapid diffusion
means that its Brownian motions
are sufficiently large at the
right time scales so that the
rates of actin monomer intercalation
can explain Listeria's
high speed. To
give the classic
Brownian ratchet some
molecular details, consider the
cartoon at right and its companion
animation (either Flash,
7Kb or QuickTime,
9Kb). Actin monomers
are 5.4 nm in diameter and F-actin
filaments are composed of two
intertwined protofilaments that
are staggered by half a monomer
(2.7 nm). As suggested
by the cartoon and its animation, Listeria must "wiggle" at
least 2.7 nm to allow intercalation
of actin monomers. The
magnitude of the "wiggles" are
inversely proportional
to the size of the bacterium
and to the viscosity of its environment. Because
diffusion is time-dependent,
the longer you wait, the larger
the magnitude of Brownian motions
and intercalation eventually
occurs. However, the high
speed of Listeria motility
implies that bacteria diffuse
very readily. Rapid diffusion
means that its Brownian motions
are sufficiently large at the
right time scales so that the
rates of actin monomer intercalation
can explain Listeria's
high speed.
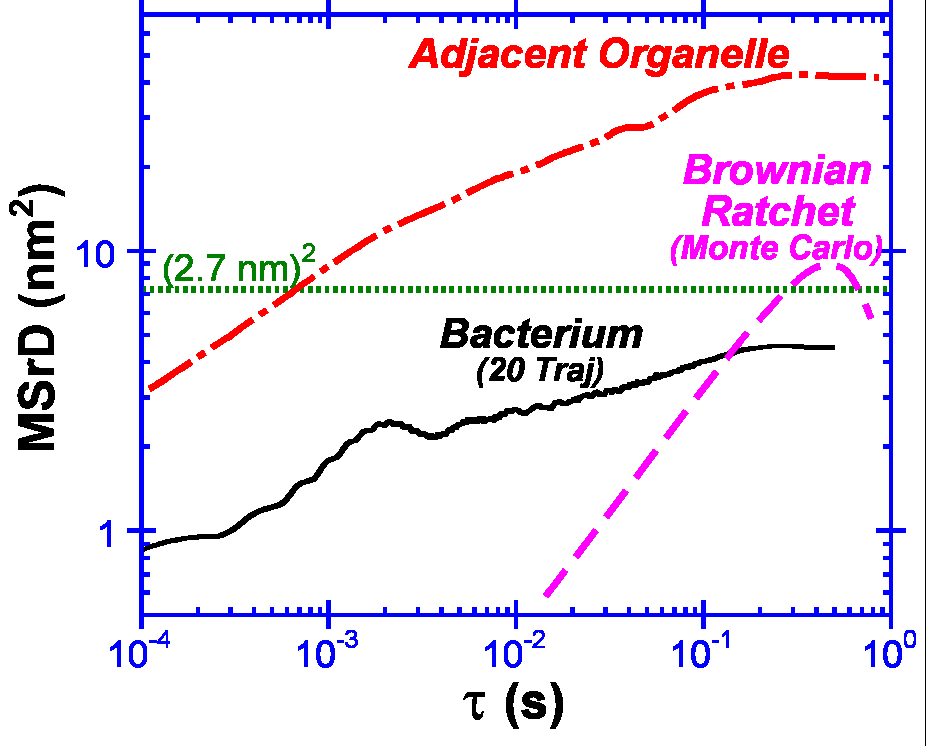 By
high resolution laser-tracking
of bacteria in living host cells,
we have shown that the classic
Brownian ratchet is not correct
for explaining Listeria motility
(Fig at left is colorized version
of Fig.4 from Kuo & McGrath
2000, Nature 407:1026-9). Not
only do Listeria "wiggle" less
than 2.7 nm, it wiggles at least
20-fold less than adjacent particles
in the cell. Hence, Listeria are
not limited by the viscosity
of the subcellular environment. They
must be binding their F-actin
tails.
By
high resolution laser-tracking
of bacteria in living host cells,
we have shown that the classic
Brownian ratchet is not correct
for explaining Listeria motility
(Fig at left is colorized version
of Fig.4 from Kuo & McGrath
2000, Nature 407:1026-9). Not
only do Listeria "wiggle" less
than 2.7 nm, it wiggles at least
20-fold less than adjacent particles
in the cell. Hence, Listeria are
not limited by the viscosity
of the subcellular environment. They
must be binding their F-actin
tails.
 Microscopically,
filaments readily bend so that
bacteria do not have to "wiggle" to
allow actin intercalation; the
filaments can do almost all the "wiggling". Often
called the "elastic
Brownian ratchet",
George Oster and colleagues (Mogilner & Oster
1996, Biophys J 71:
3030-3045) developed an alternative
to their original Brownian ratchet
model. Exaggerated in the
cartoon at right and its companion
animation (either Flash,
12Kb or QuickTime,
14Kb), the elastic Brownian
ratchet considers the bending
flexibility of filamentous actin. With
little diffusion of the bacteria,
most of the Brownian motion occurs
in the flexing of F-actin. If
filament tips flex sufficiently
far from the bacterial surface,
actin monomers can intercalate. Despite
its statistical flexing, the
longer filament applies increased
pressure to the bacterial surface. Increased
pressure will eventually cause
bacteria to move. Microscopically,
filaments readily bend so that
bacteria do not have to "wiggle" to
allow actin intercalation; the
filaments can do almost all the "wiggling". Often
called the "elastic
Brownian ratchet",
George Oster and colleagues (Mogilner & Oster
1996, Biophys J 71:
3030-3045) developed an alternative
to their original Brownian ratchet
model. Exaggerated in the
cartoon at right and its companion
animation (either Flash,
12Kb or QuickTime,
14Kb), the elastic Brownian
ratchet considers the bending
flexibility of filamentous actin. With
little diffusion of the bacteria,
most of the Brownian motion occurs
in the flexing of F-actin. If
filament tips flex sufficiently
far from the bacterial surface,
actin monomers can intercalate. Despite
its statistical flexing, the
longer filament applies increased
pressure to the bacterial surface. Increased
pressure will eventually cause
bacteria to move.
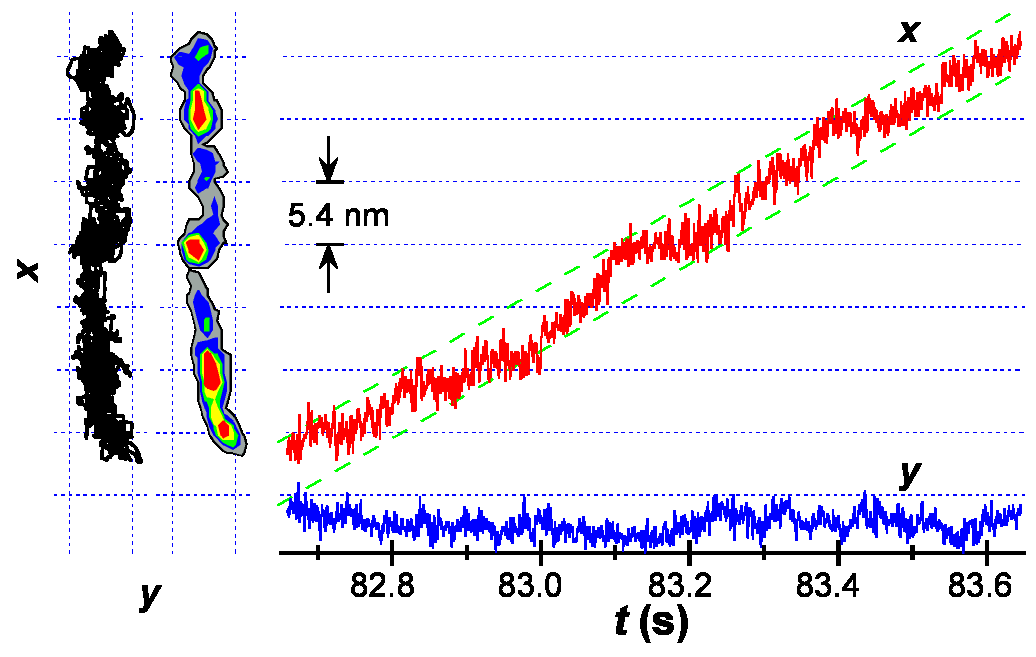 Listeria moves
with monomer-sized steps (~5.4
nm) By
high-resolution laser-tracking
of bacteria in living host cells,
we made a startling discovery. Bacteria
often paused during actin-based
motility, and pauses were spaced
the size of actin monomers (5.4
nm). The figure at right
(colorized version of Fig. 2 from Kuo & McGrath
2000, Nature 407:1026-9)
shows the "hotspots" in
pausing. The left side
of the figure shows both the
trajectory of the moving bacterium
and the "hotspots" of
bacterial position next to the
trajectory. When plotted
against time, pauses are apparent,
but some of the pauses that were
visible in the xy trajectory
become lost in the noise. Listeria moves
with monomer-sized steps (~5.4
nm) By
high-resolution laser-tracking
of bacteria in living host cells,
we made a startling discovery. Bacteria
often paused during actin-based
motility, and pauses were spaced
the size of actin monomers (5.4
nm). The figure at right
(colorized version of Fig. 2 from Kuo & McGrath
2000, Nature 407:1026-9)
shows the "hotspots" in
pausing. The left side
of the figure shows both the
trajectory of the moving bacterium
and the "hotspots" of
bacterial position next to the
trajectory. When plotted
against time, pauses are apparent,
but some of the pauses that were
visible in the xy trajectory
become lost in the noise.
Unlike
molecular motors that step (e.g.
kinesin, Svoboda et al. 1993, Nature 365:721-7), Listeria do
not move with steps of rigorously
constant sizes. Limited
by the noise in our equipment,
we can see half-monomer-sized
steps (~3 nm) and episodes
with no obvious pauses during
motility (see Supplemental Information
of Kuo & McGrath
2000, Nature 407:1026-9). On
average, steps are monomer-sized
(~5.4 nm) despite the variability
of individual intervals.
We believe
that monomer-sized steps will
prove to be general. In
preliminary studies, we've seen
monomer-sized steps in the actin-based
motility of Shigella flexneri when
infecting living host cells. We've
also seen monomer-sized steps
when reconstituting Listeria motility
in a 'test tube' using tissue
extracts to provide necessary
host factors.
How might monomer-sized
steps arise? Although actin
monomers are staggered at the
end of filaments (see cartoon),
monomers are spaced ~5.4 nm
along the sides of F-actin filaments. Progress
of a side-binding protein would
generate the appropriate spacing. However,
deeper thought reveals that we
shouldn't have seen any steps.
 Why
stepping is surprising Although
binding to the sides of filaments
could generate appropriate sized
steps, steps should not be visible
for two reasons. The first reason
is that an actin filament is
too floppy. This floppiness
is the fundamental mechanism
that allows the elastic
Brownian ratchet to
generate a gap large enough for
monomers to insert between the
ends of the filaments and the
bacterium. If a filament
were floppy enough to allow a
monomer to insert, then the filament
is not rigid enough to reveal
monomer-sized steps -- wiggles
would be as big as steps. Despite
monomer-spaced distribution of
actin along the sides of filaments,
a single filament makes a very
poor template for "stepping". Using
exaggerated flexibility, the
animation (either Flash,
27Kb or QuickTime,
29Kb) show how the flexibility
would largely obscure steps. Why
stepping is surprising Although
binding to the sides of filaments
could generate appropriate sized
steps, steps should not be visible
for two reasons. The first reason
is that an actin filament is
too floppy. This floppiness
is the fundamental mechanism
that allows the elastic
Brownian ratchet to
generate a gap large enough for
monomers to insert between the
ends of the filaments and the
bacterium. If a filament
were floppy enough to allow a
monomer to insert, then the filament
is not rigid enough to reveal
monomer-sized steps -- wiggles
would be as big as steps. Despite
monomer-spaced distribution of
actin along the sides of filaments,
a single filament makes a very
poor template for "stepping". Using
exaggerated flexibility, the
animation (either Flash,
27Kb or QuickTime,
29Kb) show how the flexibility
would largely obscure steps.
 The
second reason is that hundreds
of actin filaments form the "rocket-like" tail
that pushes bacteria. Stepping
motion implies a molecular coordination,
either filaments are molecularly
aligned or filaments are growing
in a molecularly coordinated
fashion. Without some mechanism
to impose molecular coordination, The
second reason is that hundreds
of actin filaments form the "rocket-like" tail
that pushes bacteria. Stepping
motion implies a molecular coordination,
either filaments are molecularly
aligned or filaments are growing
in a molecularly coordinated
fashion. Without some mechanism
to impose molecular coordination,
 Hypothesis
to explain appearance of steps To
generate monomer-sized steps,
we need to suppress the intrinsic
floppiness of actin filaments
illustrated above. Like
stretching a piece of string,
stretching an actin filament
will make it less floppy. With
tension, filaments become "stiffer" and
the fluctuations of a single
filament drops considerably. A
taut filament becomes an appropriate 'stepping
template' to reveal monomer-sized
steps. The animation (either Flash,
10Kb or QuickTime,
12Kb) shows how an individual
stretched filament serves as
an excellent template.
Hypothesis
to explain appearance of steps To
generate monomer-sized steps,
we need to suppress the intrinsic
floppiness of actin filaments
illustrated above. Like
stretching a piece of string,
stretching an actin filament
will make it less floppy. With
tension, filaments become "stiffer" and
the fluctuations of a single
filament drops considerably. A
taut filament becomes an appropriate 'stepping
template' to reveal monomer-sized
steps. The animation (either Flash,
10Kb or QuickTime,
12Kb) shows how an individual
stretched filament serves as
an excellent template.

To
generate a 'template' filament
within the "rocket-like" tail,
our hypothesis is that the multiple
actin filaments of the tail are
unlikely to grow at the same
rates. Indeed, the slowest
growing filaments must be stretched
taut by the other, faster growing
filaments. The animation
(either Flash,
213Kb or QuickTime,
214Kb) shows the template
filament (red) being stretched
by other filaments (purple). Consistent
with our thoughts about a binding
protein, some of the "compressed" filaments
(purple) are bound to the surface
of bacteria, but some filaments
may have released from the binding
proteins and might elongate according
to the elastic Brownian ratchet
model. The motions of binding
proteins along the sides of this
slowest-growing (red) template
filament will generate monomer-spaced
steps of the bacterium. Statistically,
a filament is 'template' for
only a limited number of steps. Another
filament becomes slowest-growing
and takes over the role as template
filament.
The cumulative action
of many filaments pushing and
a few filaments stretched taut
makes the attachment of bacteria
to their tails extremely strong. The
strength of this cumulative attachment
is revealed by the magnitude
of the fluctuations of the bacterium. Although
the 'wiggles' are smaller than
directly detectable by our laser-tracking
device, we can make an estimate
that the 'wiggles' are <0.1nm
(most chemical bonds are ~0.14nm)
and correspond to >220pN to
generate a 5.4nm step. The
value of >220pN is particularly
noteworthy because actin filaments break when
forces exceed ~400pN (Tsuda et
al. 1996, Proc Nat'l Acad
Sci, USA 93:12937-42). If
our hypothesis is correct, forces
near breaking levels are applied
to the template filament!
Implications
of monomer-sized steps There
are two major implications of
monomer-sized stepping. First,
there must be an extremely tight
binding complex that binds to
the sides of actin filaments,
and this complex can release
and re-bind to filament at monomer-sized
distances. Without a binding
complex and taut filaments (see hypothesis above),
bacteria would 'wiggle' >25-fold
more and obscure any hint of
monomer-sized steps. Second,
biophysical models must be reconsidered. The
'classic'
Brownian ratchet model must
be discarded for describing Listeria motility. However,
the physics behind the elastic
Brownian ratchet model must
occur, but the published calculations
assume that the ends of filaments
are free to 'wiggle' to allow
intercalation (Mogilner & Oster
1996, Biophys J 71:
3030-3045). If the ends
of filaments are bound by a binding
complex, the filaments must be
longer and floppier for the model
to work.
These
implications are only two of many more
implications. For example, a tight
binding complex suggests additional mechanisms
for regulating cellular protrusions generated
by actin polymerization (see examples). The
more points of regulation that we understand,
the more likely we can control protrusive
processes, both in clinical and engineering
applications. Only additional research
can determine if these more speculative
ideas are correct.
|
