





How do remodelers sense DNA outside the nucleosome?
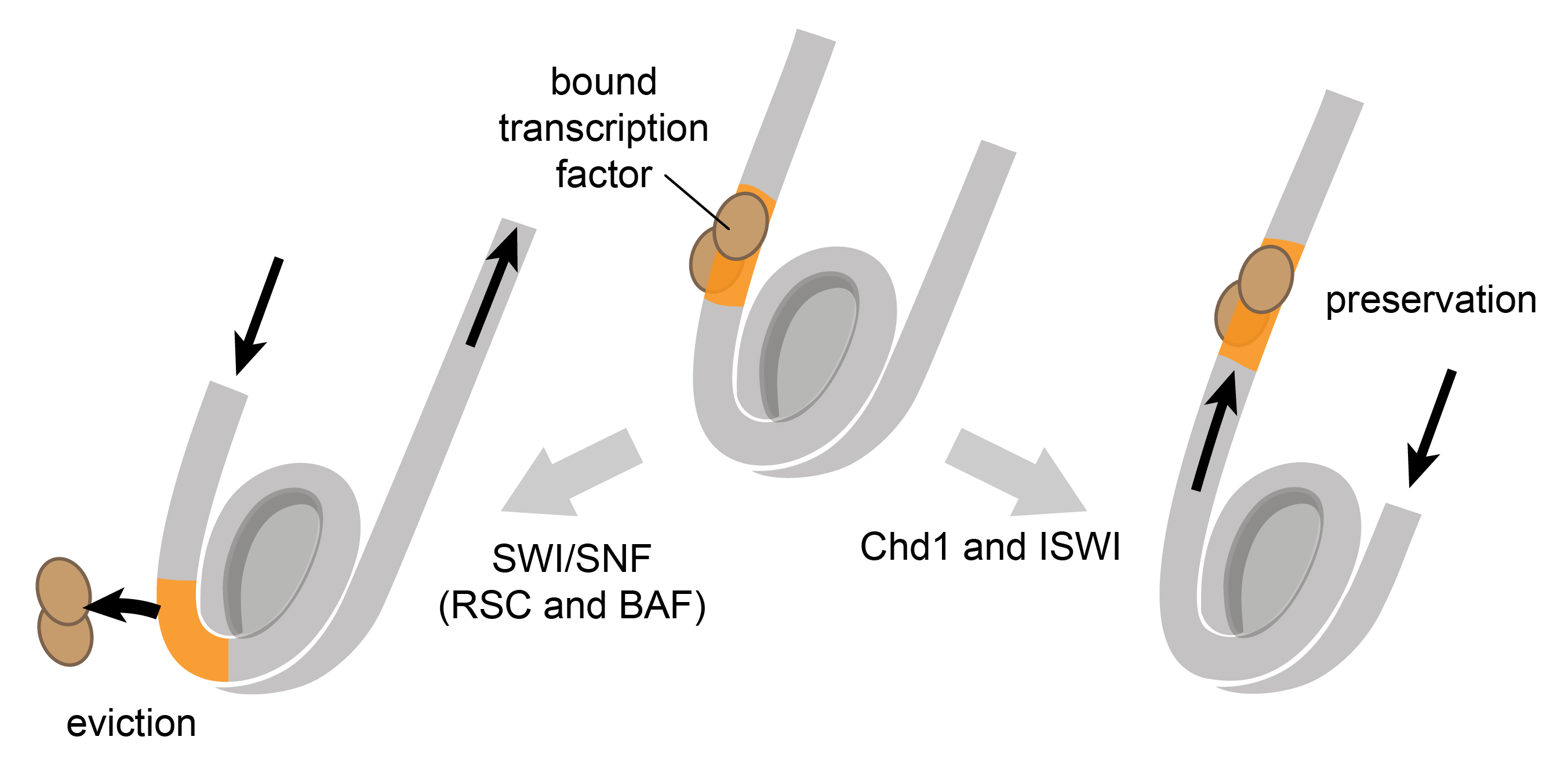
How do remodelers generate particular patterns of nucleosomes? The ability of remodelers to generate evenly-spaced arrays (Chd1 and ISWI) or disrupt those arrays (SWI/SNF) appears to arise from sensitivity to DNA flanking the nucleosome. SWI/SNF, which has long been known to be a transcriptional activator for disrupting chromatin at promoters, can slide nucleosomes on top of transcription factors, which causes those factors to dissociate (Nagaich et al., 2004; Li et al., 2015). It can also slide nucleosomes into neighboring nucleosomes, which is believed to stimulate histone ejection (Dechassa et al., 2010).
In contrast, Chd1 and ISWI remodelers generally require DNA to be available on the "entry" side in order to reposition or slide nucleosomes (Yang et al., 2006; Stockdale et al., 2006). In addition to creating evenly-spaced arrays (Lusser et al., 2005), ISWI and Chd1 also preferentially shift nucleosomes away from bound transcription factors (Kang et al., 2002; Li et al., 2015; Nodelman et al., 2016), consistent with a sensitivity to DNA flanking the nucleosome.
 |
We are interested in how remodelers sense DNA outside the nucleosome core. For Chd1, we showed that DNA sensing relies on the DNA-binding domain. The DNA-binding domain, however, is not essential for nucleosome sliding, as we found it could be effectively replaced by foreign binding domain (McKnight et al., 2011; Patel et al., 2013; Nodelman & Bowman, 2013; McKnight et al., 2016). Thus, the DNA-binding domain appears to have a regulatory rather than a mechanical role in the process.
Recently, we proposed that DNA sensing by Chd1 could be achieved through direct domain-domain communication (Nodelman et al., 2017). Consistent with cryo-EM structures (Farnung et al., 2017; Sundaramoorthy et al., 2018), we found that all three Chd1 domains cluster together on the nucleosome. We previously showed that the Chd1 chromodomains and ATPase can pack closely together, with the chromodomains blocking a key DNA-binding surface of the ATPase motor (Hauk et al., 2010). When clustered on the nucleosome, the chromodomains and ATPase maintain a close association, though the chromodomains are slightly displaced to allow the ATPase to fully engage with nucleosomal DNA. Packing against this chromo-ATPase unit, the DNA-binding domain sits on DNA poised to exit the nucleosome. Based on this clustering of domains, we have proposed an inhibitory model for sensing DNA (Nodelman et al., 2017).
 |
We are currently exploring whether and how the DNA-binding domain may be communicating with the ATPase motor. One hypothesis is that the DNA-binding domain affects the ATPase via the chromodomains, which appear poised to inhibit the remodeler. An open question is how such a domain arrangement may also make the remodeler sensitive to flanking DNA on the other (entry) side of the nucleosome. Chd1 also has a regulatory element on the C-terminal side of the ATPase motor, called the Bridge (Hauk et al., 2010). This element, which we predicted for ISWI, was also shown to be important for sensing flanking DNA (Clapier et al., 2012; Leonard & Narlikar, 2015). How this element contributes to DNA sensing in the wildtype enzyme remains unclear.
| back to Research Overview |