






Research interests in the Bowman lab
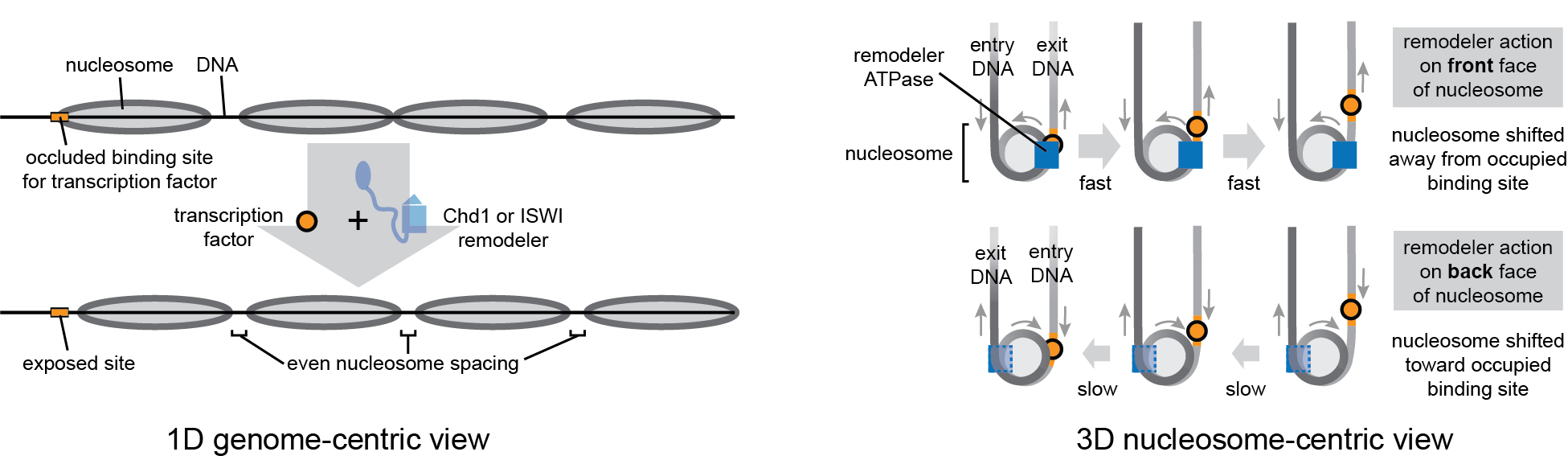
The vast majority of eukaryotic genomes are organized into regularly spaced nucleosomal arrays, which prevent spurious transcription initiation at cryptic promoters. Regularly spaced arrays are primarily established and maintained by Chd1- and ISWI-type chromatin remodelers. Although nucleosomes can be moved back-and-forth, remodelers slide nucleosomes unidirectionally, determined by the face of the nucleosome disk where the remodeler ATPase acts. Due to the intrinsic two-fold symmetry of the histone octamer, remodelers typically act at either of two internal sites (called superhelix 2, or SHL2), on opposites faces of the nucleosome. Chd1 and ISWI remodelers preferentially shift nucleosomes away from bound transcription factors and this biased directionality appears to result from stimulating remodelers on exit DNA and slowing them down on entry. A fundamental question is how remodelers integrate information about their environment to produce particular remodeling outcomes.
Research interests in the Bowman lab
 |
An exciting breakthrough in understanding nucleosome sliding was the discovery that the remodeler ATPase initiates DNA movement without ATP (Winger et al., 2017). By pulling DNA towards itself on one side, the remodeler ATPase locally changes the twist of DNA, which effectively absorbs a base pair. This DNA distortion occurs in ADP and nucleotide-free conditions, when the ATPase is in an open state. Upon binding to ATP, the ATPase closes, which reverses the DNA distortion, pushing the extra base pair further onto the nucleosome (Nodelman & Bowman). In collaboration with Sebastian Deindl’s group, we showed that these shifts occur sequentially, with DNA first pulled onto the nucleosome before being pushed out the exit side (Sebantsev, Levendosky et al., 2019). More recent work with Jean-Paul Armache’s group has revealed that the distorted DNA intermediate has an A-form-like geometry, where one strand is locally shifted ahead of the other (Nodelman, Das et al., 2022).
 |
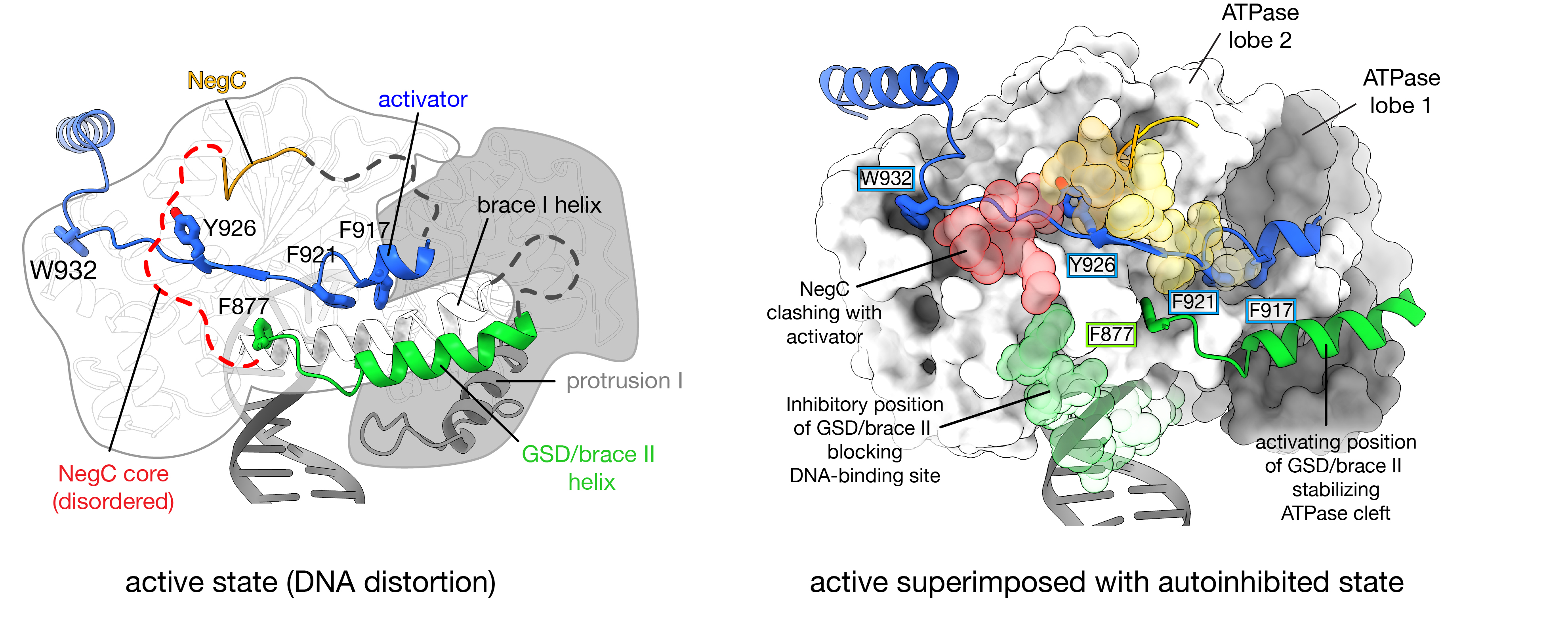
Also in collaboration with the Armache group, we have discovered that the DNA distortion stage appears to be a regulatory checkpoint (Nodelman, Folkwein et al., 2025). As shown by cryo-EM and biochemistry, we identified two elements (GSD/brace II helix and brace stabilizer) that bind to the central cleft of the ATPase motor and help stabilize the ATPase open conformation needed for DNA distortion. Interestingly, immediately between these positive elements is a negative regulatory segment(NegC) that directly competes for binding to the ATPase motor. We first observed the path of NegC in our Chd1 crystal structure (Hauk, McKnight et al., 2010 ), which sterically conflicts with the path of the brace stabilizer. NegC binding to the ATPase would also prevent the GSD/brace II helix from reaching the ATPase central cleft. We propose that the location of the DNA-binding domain tips the balance in favor of either activators or the NegC inhibitor.
 |
Recently, the groups of Zhucheng Chen and Lucas Farnung captured remodeler ATPases in inhibited states on the nucleosome (Sia et al., 2025; James & Farnung, 2025 ). These structures confirm our predictions about partially disengaged states of the ATPase on the nucleosome (Nodelman et al., 2021) and raise questions of remodeler domain dynamics in different contexts. Ultimately, we hope to reveal deep mechanistic insights into remodeler action, remodeler regulation, and nucleosome behaviors during chromatin remodeling.